青藏高原被誉为地球的“第三极”,这里气候极端,但同时是生物多样性的宝库。全球约一半的授粉昆虫熊蜂物种(bumblebee)都栖息于此,因此青藏高原及周边区域也被称熊蜂的“起源中心”和“演化摇篮”。然而,一个谜团始终困扰着科学家:这些熊蜂是如何从高寒的“世界屋脊”扩散到周边乃至更远地区的低地环境,并成功适应巨大温差的?
近期,一项由河北大学生命科学学院陈兵教授团队牵头开展的联合研究,为我们揭开了这一演化谜题的重要一角。研究团队以广泛分布于中国的火红熊蜂(Bombus pyrosoma)为模型,综合运用基因组学、生理学和行为学手段,发现染色体倒位这种特殊的基因组结构变异,在熊蜂适应不同海拔环境并成功向外扩散的过程中,扮演了至关重要的“引擎”角色,而且这一基因组变异规律在另外22个熊蜂中普遍存在。该成果发表于《美国科学院院刊》(PNAS),由生命科学学院博士生姜春艳与讲师岳雷为共同第一作者、陈兵教授与中国农业科学院蜜蜂研究所黄家兴研究员为共同通讯作者。

一、 追溯迁徙之路:从高原到低地的清晰足迹
研究者们首先对覆盖其全分布区的119只火红熊蜂进行全基因组测序,成功重建了该物种的迁徙历史。系统发育分析显示,现生的低地种群均源自青藏高原的高地祖先种群。大约在95万年前,一次显著的分化发生了,种群在约海拔2000米处“分道扬镳”,形成了遗传结构显著不同的高地和低地谱系。基因流分析进一步证实,两者之间的基因交流非常有限,这为独立的局部适应创造了条件。
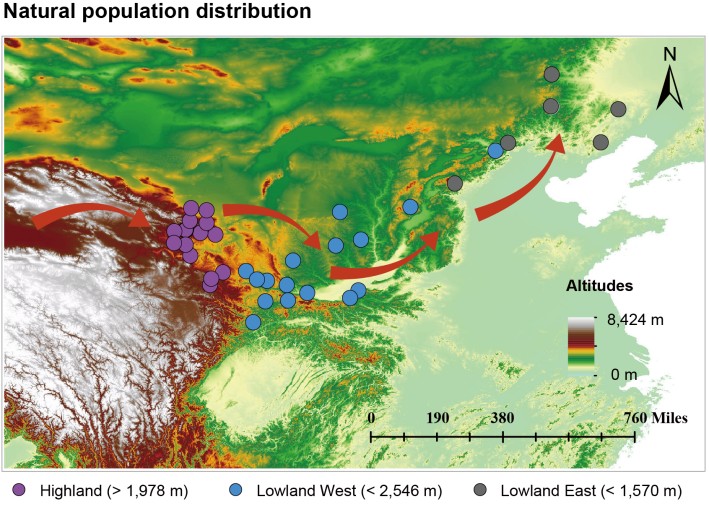
火红熊蜂的高原起源与迁徙路线
二、 锁定关键机制:染色体倒位的“海拔开关”
究竟是基因组什么遗传变化驱动了这种快速的海拔适应?通过全基因组多种分析,研究团队聚焦到了染色体倒位——一种染色体片段发生180度旋转重排的结构变异。他们在火红熊蜂基因组中鉴定出四个主要的倒位区域。令人惊讶的是,这些倒位的频率呈现出清晰的海拔梯度:在低地种群中,倒位类型高度流行(>73.7%为纯合倒位),而在高海拔种群中,则主要以祖先的非倒位类型为主。频率的急剧变化恰恰发生在海拔2000米附近,与种群遗传分界线完美吻合。
三、 表型与功能的印证:冷暖各有所长的生存策略
基因组的变化最终要体现在生物体的功能上。通过形态变异分析发现,高海拔熊蜂工蜂体型相对更小,但翅膀相对更大,这有助于在空气稀薄的环境中飞行。关键的差异体现在温度适应上:高地熊蜂是“抗寒高手”,在寒冷环境下能保持更高的胸温、更低的冻僵率,并维持正常的飞行能力和觅食积极性;而低地熊蜂则是“耐热专家”,在高温下拥有更高的临界热最大值和更稳定的飞行表现。这种“冷暖专精”的差异,正是它们在不同栖境生存下来的秘诀。
四、 基因解码:elovl6如何成为“寒冷飞行”的关键
那么,倒位区域内有哪些基因在起作用?群体遗传学分析锁定了大量正选择基因,其中41%都位于倒位区内,它们显著富集于脂肪酸代谢等通路。一个名为 elovl6 的基因脱颖而出。该基因编码的酶负责催化长链脂肪酸(如硬脂酸C18:0)的合成。研究发现,高海拔熊蜂的飞行肌肉中,elovl6表达水平更高,硬脂酸含量也更丰富。当给熊蜂补充硬脂酸后,它们在寒冷环境下的飞行能力得到了显著提升!这证明,倒位可能通过“锁住”包括elovl6在内的有利基因组合,促进了高海拔熊蜂对寒冷飞行代谢的适应。
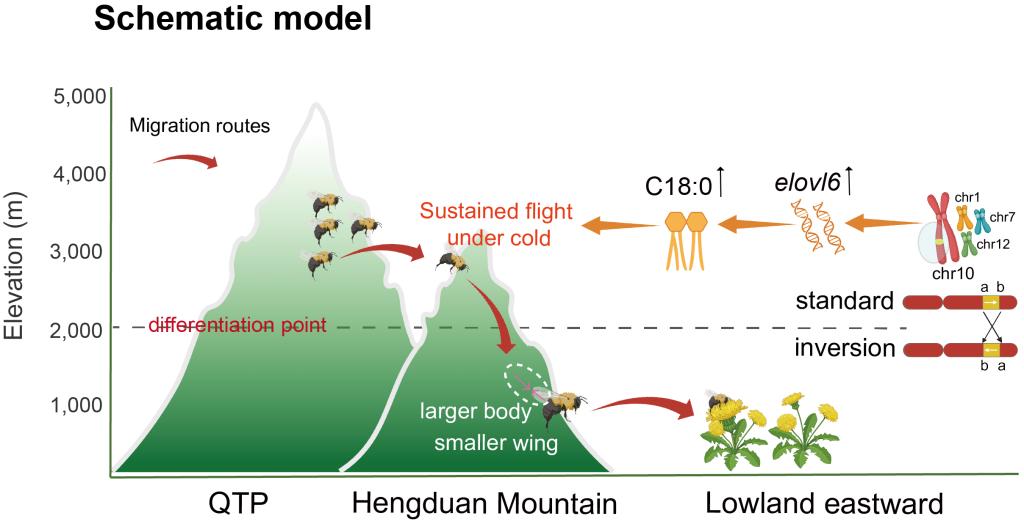
染色体倒位促进火红熊蜂的海拔适应及走出青藏高原
五、 普遍的意义:染色体倒位是熊蜂海拔多样化的普遍推手
这一现象是个例吗?为了回答这个问题,研究者比较了包括17个青藏高原土著种在内的22个熊蜂物种。其中从头组装了11个本地熊蜂染色体级别的基因组。分析发现,染色体倒位在熊蜂中广泛存在,且其数量在分布于不同海拔的姐妹物种间显著更多。特别是其中一个在火红熊蜂中关键的染色体10倒位,也出现在其他多个低地熊蜂物种中。这表明,染色体倒位很可能是熊蜂乃至其他高山生物适应不同海拔环境、实现多样化的一个普遍且重要的进化机制。
这项研究不仅首次系统阐明了染色体倒位在熊蜂“走出青藏高原”过程中的核心作用,揭示了从基因组结构变异到生理行为适应的完整链条,也为理解全球高山生物多样性的形成提供了宝贵的新见解。随着气候变化,高山物种的生存面临挑战,而理解它们适应环境的遗传“工具箱”,对于未来的生物多样性保护至关重要。
信息来源:
Jiang et al. Ancestral and local adaptation contribute to dispersal out of the Qinghai–Tibet Plateau in a bumblebee. Proceedings of the National Academy of Sciences, 2026, 123 (6), e2513080122. DOI:https://doi.org/10.1073/pnas.2513080122








